Gray's Anatomy:
|
Du er her: Hjem >> BiologiskAntropologi >> Osteologi >> GraysAnatomy >> Osteology_introduction.php
II. Osteology
Introduction
|
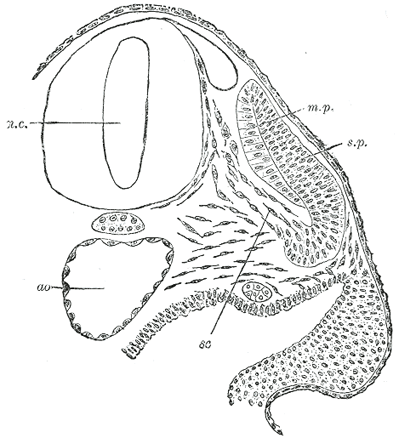
FIG. 64– Transverse section of a human embryo of the third week to show the differentiation of the primitive segment. (Kollmann.) ao. Aorta. m.p. Muscle-plate. n.c. Neural canal. sc. Sclerotome. s.p. cutis-plate. | |
| THE GENERAL framework of the body is built up mainly of a series of bones, supplemented, however, in certain regions by pieces of cartilage; the bony part of the framework constitutes the skeleton. |
| In the skeleton of the adult there are 206 distinct bones, as follows:— |
Axial
Skeleton |
Vertebral column |
26 |
|
| Skull |
22 |
| Hyoid bone |
1 |
| Ribs and sternum |
25 |
| |
|
— |
74 |
Appendicular
Skeleton |
Upper extremities |
64 |
|
| Lower extremities |
62 |
| |
|
— |
126 |
| Auditory ossicles |
|
|
6 |
| |
|
|
— |
| |
Total |
|
206 |
|
| The patellæ are included in this enumeration, but the smaller sesamoid bones are not reckoned. |
| Bones are divisible into four classes: Long, Short, Flat, and Irregular. |
| | | Long Bones.—The long bones are found in the limbs, and each consists of a body or shaft and two extremities. The body, or diaphysis is cylindrical, with a central cavity termed the medullary canal; the wall consists of dense, compact tissue of considerable thickness in the middle part of the body, but becoming thinner toward the extremities; within the medullary canal is some cancellous tissue, scanty in the middle of the body but greater in amount toward the ends. The extremities are generally expanded, for the purposes of articulation and to afford broad surfaces for muscular attachment. They are usually developed from separate centers of ossification termed epiphyses, and consist of cancellous tissue surrounded by thin compact bone. The medullary canal and the spaces in the cancellous tissue are filled with marrow. The long bones are not straight, but curved, the curve generally taking place in two planes, thus affording greater strength to the bone. The bones belonging to this class are: the clavicle, humerus, radius, ulna, femur, tibia, fibula, metacarpals, metatarsals, and phalanges. |
| | | Short Bones.—Where a part of the skeleton is intended for strength and compactness combined with limited movement, it is constructed of a number of short bones, as in the carpus and tarsus. These consist of cancellous tissue covered by a thin crust of compact substance. The patellæ, together with the other sesamoid bones, are by some regarded as short bones. |
| | | Flat Bones.—Where the principal requirement is either extensive protection or the provision of broad surfaces for muscular attachment, the bones are expanded into broad, flat plates, as in the skull and the scapula. These bones are composed of two thin layers of compact tissue enclosing between them a variable quantity of cancellous tissue. In the cranial bones, the layers of compact tissue are familiarly known as the tables of the skull; the outer one is thick and tough; the inner is thin, dense, and brittle, and hence is termed the vitreous table. The intervening cancellous tissue is called the diploë, and this, in certain regions of the skull, becomes absorbed so as to leave spaces filled with air (air-sinuses) between the two tables. The flat bones are: the occipital, parietal, frontal, nasal, lacrimal, vomer, scapula, os coxæ (hip bone), sternum, ribs, and, according to some, the patella. |
| | | Irregular Bones.—The irregular bones are such as, from their peculiar form, cannot be grouped under the preceding heads. They consist of cancellous tissue enclosed within a thin layer of compact bone. The irregular bones are: the vertebræ, sacrum, coccyx, temporal, sphenoid, ethmoid, zygomatic, maxilla, mandible, palatine, inferior nasal concha, and hyoid. |
| | | Surfaces of Bones.—If the surface of a bone be examined, certain eminences and depressions are seen. These eminences and depressions are of two kinds: articular and non-articular. Well-marked examples of articular eminences are found in the heads of the humerus and femur; and of articular depressions in the glenoid cavity of the scapula, and the acetabulum of the hip bone. Non-articular eminences are designated according to their form. Thus, a broad, rough, uneven elevation is called a tuberosity, protuberance, or process, a small, rough prominence, a tubercle; a sharp, slender pointed eminence, a spine; a narrow, rough elevation, running some way along the surface, a ridge, crest, or line. Non-articular depressions are also of variable form, and are described as fossæ, pits, depressions, grooves, furrows, fissures, notches, etc. These non-articular eminences and depressions serve to increase the extent of surface for the attachment of ligaments and muscles, and are usually well-marked in proportion to the muscularity of the subject. A short perforation is called a foramen, a longer passage a canal. |
| |
1. Development of the Skeleton |
| |
| | | The Skeleton.—The skeleton is of mesodermal origin, and may be divided into (a) that of the trunk (axial skeleton), comprising the vertebral column, skull, ribs, and sternum, and (b) that of the limbs (appendicular skeleton). |
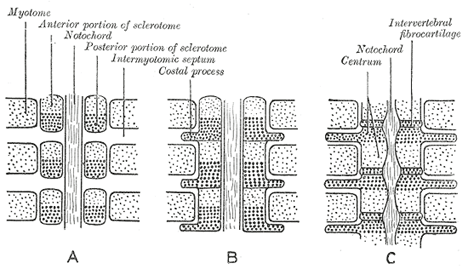
| | | The Vertebral Column.—The notochord (Fig. 19) is a temporary structure and forms a central axis, around which the segments of the vertebral column are developed. (*11 It is derived from the entoderm, and consists of a rod of cells, which lies on the ventral aspect of the neural tube and reaches from the anterior end of the mid-brain to the extremity of the tail. On either side of it is a column of paraxial mesoderm which becomes subdivided into a number of more or less cubical segments, the primitive segments (Figs. 19 and 20). These are separated from one another by intersegmental septa and are arranged symmetrically on either side of the neural tube and notochord: to every segment a spinal nerve is distributed. At first each segment contains a central cavity, the myocœl, but this is soon filled with a core of angular and spindle-shaped cells. The cells of the segment become differentiated into three groups, which form respectively the cutis-plate or dermatome, the muscle-plate or myotome, and the sclerotome (Fig. 64). The cutis-plate is placed on the lateral and dorsal aspect of the myocœl, and from it the true skin of the corresponding segment is derived; the muscle-plate is situated on the medial side of the cutis-plate and furnishes the muscles of the segment. The cells of the sclerotome are largely derived from those forming the core of the myocœl, and lie next the notochord. Fusion of the individual sclerotomes in an antero-posterior direction soon takes place, and thus a continuous strand of cells, the sclerotogenous layer, is formed along the ventro-lateral aspects of the neural tube. The cells of this layer proliferate rapidly, and extending medialward surround the notochord; at the same time they grow backward on the lateral aspects of the neural tube and eventually surround it, and thus the notochord and neural tube are enveloped by a continuous sheath of mesoderm, which is termed the membranous vertebral column. In this mesoderm the original segments are still distinguishable, but each is now differentiated into two portions, an anterior, consisting of loosely arranged cells, and a posterior, of more condensed tissue (Fig. 65, A and B). Between the two portions the rudiment of the intervertebral fibrocartilage is laid down (Fig. 65, C). Cells from the posterior mass grow into the intervals between the myotomes (Fig. 65, B and C) of the corresponding and succeeding segments, and extend both dorsally and ventrally; the dorsal extensions surround the neural tube and represent the future vertebral arch, while the ventral extend into the body-wall as the costal processes. The hinder part of the posterior mass joins the anterior mass of the succeeding segment to form the vertebral body. Each vertebral body is therefore a composite of two segments, being formed from the posterior portion of one segment and the anterior part of that immediately behind it. The vertebral and costal arches are derivatives of the posterior part of the segment in front of the intersegmental septum with which they are associated. |
| |
|
FIG. 65– Scheme showing the manner in which each vertebral centrum is developed from portions of two adjacent segments. | | |
| This stage is succeeded by that of the cartilaginous vertebral column. In the fourth week two cartilaginous centers make their appearance, one on either side of the notochord; these extend around the notochord and form the body of the cartilaginous vertebra. A second pair of cartilaginous foci appear in the lateral parts of the vertebral bow, and grow backward on either side of the neural tube to form the cartilaginous vertebral arch, and a separate cartilaginous center appears for each costal process. By the eighth week the cartilaginous arch has fused with the body, and in the fourth month the two halves of the arch are joined on the dorsal aspect of the neural tube. The spinous process is developed from the junction of the two halves of the vertebral arch. The transverse process grows out from the vertebral arch behind the costal process. |
| In the upper cervical vertebræ a band of mesodermal tissue connects the ends of the vertebral arches across the ventral surfaces of the intervertebral fibrocartilages. This is termed the hypochordal bar or brace; in all except the first it is transitory and disappears by fusing with the fibrocartilages. In the atlas, however, the entire bow persists and undergoes chondrification; it develops into the anterior arch of the bone, while the cartilage representing the body of the atlas forms the dens or odontoid process which fuses with the body of the second cervical vertebra. |
|
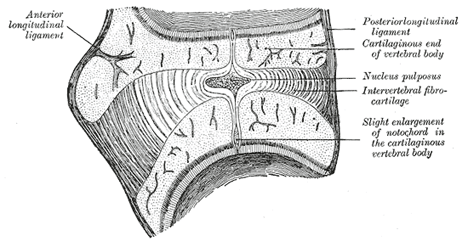
FIG. 66– Sagittal section through an intervertebral fibrocartilage and adjacent parts of two vertebræ of an advanced sheep’s embryo. (Kölliker.) | | |
| The portions of the notochord which are surrounded by the bodies of the vertebræ atrophy, and ultimately disappear, while those which lie in the centers of the intervertebral fibrocartilages undergo enlargement, and persist throughout life as the central nucleus pulposus of the fibrocartilages (Fig. 66). |
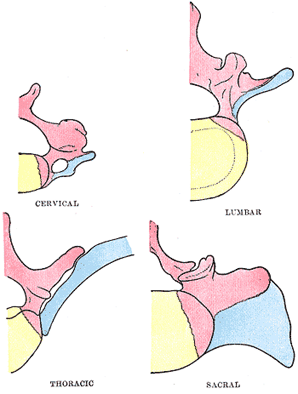
| | | The Ribs.—The ribs are formed from the ventral or costal processes of the primitive vertebral bows, the processes extending between the muscle-plates. In the thoracic region of the vertebral column the costal processes grow lateralward to form a series of arches, the primitive costal arches. As already described, the transverse process grows out behind the vertebral end of each arch. It is at first connected to the costal process by continuous mesoderm, but this becomes differentiated later to form the costotransverse ligament; between the costal process and the tip of the transverse process the costotransverse joint is formed by absorption. The costal process becomes separated from the vertebral bow by the development of the costocentral joint. In the cervical vertebrœ (Fig. 67) the transverse process forms the posterior boundary of the foramen transversarium, while the costal process corresponding to the head and neck of the rib fuses with the
body of the vertebra, and forms the antero-lateral boundary of the foramen. The distal portions of the primitive costal arches remain undeveloped; occasionally the arch of the seventh cervical vertebra undergoes greater development, and by the formation of costovertebral joints is separated off as a rib. In the lumbar region the distal portions of the primitive costal arches fail; the proximal portions fuse with the transverse processes to form the transverse processes of descriptive anatomy. Occasionally a movable rib is developed in connection with the first lumbar vertebra. In the sacral region costal processes are developed only in connection with the upper three, or it may be four, vertebræ the processes of adjacent segments fuse with one another to form the lateral parts of the sacrum. The coccygeal vertebrœ are devoid of costal processes. |
|
FIG. 67– Diagrams showing the portions of the adult vertebræ derived respectively from the bodies, vertebral arches, and costal processes of the embryonic vertebræ. The bodies are represented in yellow, the vertebral arches in red, and the costal processes in blue. | | |
| | | The Sternum.—The ventral ends of the ribs become united to one another by a longitudinal bar termed the sternal plate, and opposite the first seven pairs of ribs these sternal plates fuse in the middle line to form the manubrium and body of the sternum. The xiphoid process is formed by a backward extension of the sternal plates. |
| | | The Skull.—Up to a certain stage the development of the skull corresponds with that of the vertebral column; but it is modified later in association with the expansion of the brain-vesicles, the formation of the organs of smell, sight, and hearing, and the development of the mouth and pharynx. |
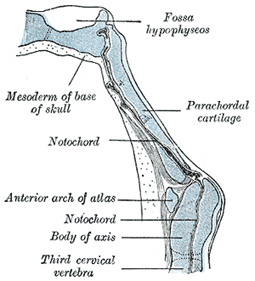
| The notochord extends as far forward as the anterior end of the mid-brain, and becomes partly surrounded by mesoderm (Fig. 68). The posterior part of this mesodermal investment corresponds with the basilar part of the occipital bone, and shows a subdivision into four segments, which are separated by the roots of the hypoglossal nerve. The mesoderm then extends over the brain-vesicles, and thus the entire brain is enclosed by a mesodermal investment, which is termed the membranous cranium. From the inner layer of this the bones of the skull and the membranes of the brain are developed; from the outer layer the muscles, bloodvessels, true skin, and subcutaneous tissues of the scalp. In the shark and dog-fish this membranous cranium undergoes complete chondrification, and forms the cartilaginous skull or chondrocranium of these animals. In mammals, on the other hand, the process of chondrification is limited to the base of the skull—the roof and sides being covered in by membrane. Thus the bones of the base of the skull are preceded by cartilage, those of the roof and sides by membrane. The posterior part of the base of the skull is developed around the notochord, and exhibits a segmented condition analogous to that of the vertebral column, while the anterior part arises in front of the notochord and shows no regular segmentation. The base of the skull may therefore be divided into (a) a chordal or vertebral, and (b) a prechordal or prevertebral portion. |
|
FIG. 68– Sagittal section of cephalic end of notochord. (Keibel.) | | |
|
FIG. 69– Diagrams of the cartilaginous cranium. (Wiedersheim.) | | |
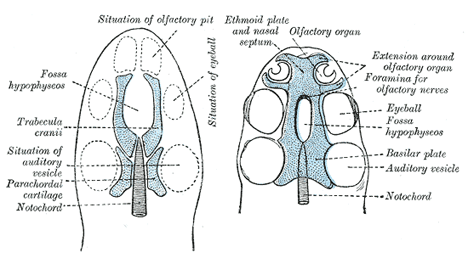
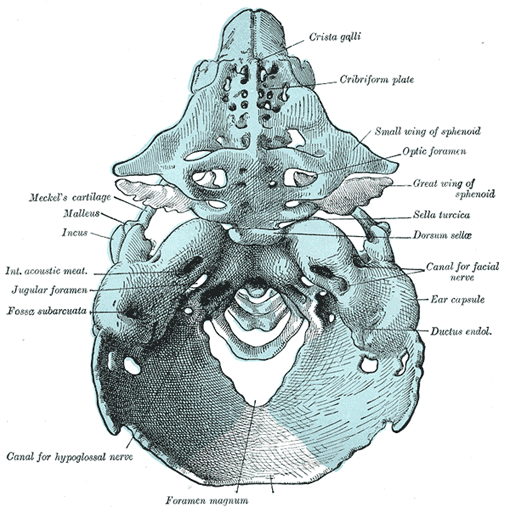
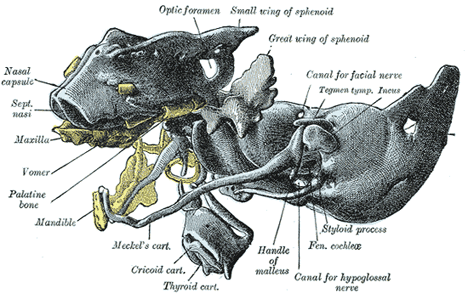
| In the lower vertebrates two pairs of cartilages are developed, viz., a pair of parachordal cartilages, one on either side of the notochord; and a pair of prechordal cartilages, the trabeculæ cranii, in front of the notochord (Fig. 66). The parachordal cartilages (Fig. 69) unite to form a basilar plate, from which the cartilaginous part of the occipital bone and the basi-sphenoid are developed. On the lateral aspects of the parachordal cartilages the auditory vesicles are situated, and the mesoderm enclosing them is soon converted into cartilage, forming the cartilaginous ear-capsules. These cartilaginous ear-capsules, which are of an oval shape, fuse with the sides of the basilar plate, and from them arise the petrous and mastoid portions of the temporal bones. The trabeculæ cranii (Fig. 69) are two curved bars of cartilage which embrace the hypophysis cerebri; their posterior ends soon unite with the basilar plate, while their anterior ends join to form the ethmoidal plate, which extends forward between the fore-brain and the olfactory pits. Later the trabeculæ meet and fuse below the hypophysis, forming the floor of the fossa hypophyseos and so cutting off the anterior lobe of the hypophysis from the stomodeum. The median part of the ethmoidal plate forms the bony and cartilaginous parts of the nasal septum. From the lateral margins of the trabeculæ cranii three processes grow out on either side. The anterior forms the ethmoidal labyrinth and the lateral and alar cartilages of the nose; the middle gives rise to the small wing of the sphenoid, while from the posterior the great wing and lateral pterygoid plate of the sphenoid are developed (Figs. 70, 71). The bones of the vault are of membranous formation, and are termed dermal or covering bones. They are partly developed from the mesoderm of the membranous cranium, and partly from that which lies outside the entoderm of the foregut. They comprise the upper part of the occipital squama (interparietal), the squamæ and tympanic parts of the temporals, the parietals, the frontal, the vomer, the medial pterygoid plates, and the bones of the face. Some of them remain distinct throughout life, e.g., parietal and frontal, while others join with the bones of the chondrocranium, e.g., interparietal, squamæ of temporals, and medial pterygoid plates. |
|
FIG. 70– Model of the chondrocranium of a human embryo, 8 cm. long. (Hertwig.) The membrane bones are not represented. | | |
| Recent observations have shown that, in mammals, the basi-cranial cartilage, both in the chordal and prechordal regions of the base of the skull, is developed as a single plate which extends from behind forward. In man, however, its posterior part shows an indication of being developed from two chondrifying centers which fuse rapidly in front and below. The anterior and posterior thirds of the cartilage surround the notochord, but its middle third lies on the dorsal aspect of the notochord, which in this region is placed between the cartilage and the wall of the pharynx. |
|
FIG. 71– The same model as shown in Fig. 70 from the left side. Certain of the membrane bones of the right side are represented in yellow. (Hertwig.) | | |
|
Note 11. In the amphioxus the notochord persists and forms the only representative of a skeleton in that animal. [back]
|
|
|